Содержание

Клеточная стенка — оболочка клетки, расположенная снаружи от цитоплазматической мембраны и выполняющая структурные, защитные и транспортные функции. Обнаруживается у большинства бактерий, архей, грибов и растений. Животные и многие простейшие не имеют клеточной стенки.
Содержание
Клеточные стенки прокариот [ править | править код ]
Клеточные стенки бактерий состоят из пептидогликана (муреина) и бывают двух типов: грамположительного и грамотрицательного. Клеточная стенка грамположительного типа состоит исключительно из толстого слоя пептидогликана, плотно прилегающего к клеточной мембране и пронизанного тейхоевыми и липотейхоевыми кислотами. При грамотрицательном типе слой пептидогликана существенно тоньше, между ним и плазматической мембраной находится периплазматическое пространство, а снаружи клетка окружена ещё одной мембраной, представленной т. н. липополисахаридом и являющаяся пирогенным эндотоксином грамотрицательных бактерий.
Клеточные стенки грибов [ править | править код ]
Клеточные стенки грибов состоят из хитина и глюканов.
Клеточные стенки водорослей [ править | править код ]
Большинство водорослей имеют клеточную стенку из целлюлозы и различных гликопротеинов. Включения дополнительных полисахаридов имеют большое таксономическое значение.
Диатомовые водоросли синтезируют свою клеточную стенку из кремнезёма.
Клеточные стенки высших растений [ править | править код ]
Важнейшей отличительной особенностью растительной клетки является наличие прочной клеточной стенки, основным компонентом которой является целлюлоза. Клеточная стенка высших растений представляет собой сложноорганизованный, преимущественно полимерный, внеклеточный матрикс, окружающий каждую клетку. Растительная клетка, лишённая клеточной стенки, обозначается термином протопласт. В клеточных стенках растений существуют углубления — поры, через которые проходят цитоплазматические канальца — плазмодесмы, осуществляющие контакт соседних клеток и обмен веществами между ними.
Химический состав и пространственная организация полимеров клеточной стенки отличаются у разных видов, клеток разных тканей одного растения и иногда у разных частей стенки вокруг одного протопласта.
Кроме того, строение клеточной стенки изменяется в онтогенезе растительного организма. Первичная клеточная стенка формируется при делении и сохраняется во время роста клетки. Формирование вторичной клеточной стенки происходит с внутренней стороны от первичной стенки и связано с окончанием роста и специализацией (дифференцировкой) клеток растения. Снаружи от первичной клеточной стенки, между первичными стенками двух соседних клеток, располагается срединная пластинка (состоит преимущественно из кальциевых и магниевых солей пектиновых веществ).

Первичная клеточная стенка высших растений состоит из трёх взаимодействующих, но структурно независимых трехмерных сетей полимеров. Основная сеть состоит из фибрилл целлюлозы и связывающих их гемицеллюлоз (или сшивочных гликанов). Вторая сеть состоит из пектиновых веществ. Третья сеть представлена, как правило, структурными белками клеточной стенки. Также следует отметить, что у растений клады commelinids (группа в системах APG) и у представителей семейства Маревые первичная клеточная стенка содержит значительное количество ароматических веществ (гидроксикоричные кислоты, главным образом феруловая и п-кумаровая). При этом у представителей клады commelinids гидроксикоричные кислоты присоединяются к сшивочным гликанам (к глюкуроноарабиноксиланам), а у семейства Маревые к пектиновым веществам (к рамногалактуронанам I).
Растительные клеточные стенки выполняют целый ряд функций: они обеспечивают жёсткость клетки для структурной и механической поддержки, придают форму клетке, направление её роста и в конечном счете морфологию всему растению. Клеточная стенка также противодействует тургору, то есть осмотическому давлению, когда дополнительное количество воды поступает в растения. Клеточные стенки защищают от патогенов, проникающих из окружающей среды, и запасают углеводы для растения.
В отличие от животных и многих простейших, у растений, бактерий и грибов, почти все клетки имеют стенку, лежащую кнаружи от цитоплазматической мембраны и обладающую повышенной прочностью. Основная функция данной структуры — опора и защита.
Клеточные стенки (или клеточные оболочки) строятся из веществ, синтезируемых самими клетками. Их химический состав различен у растений, грибов и прокариот. Кроме того, даже у одного растения у различных клеток состав стенок несколько различен.
Клеточная стенка растений состоит в основном из целлюлозы. Целлюлоза — это полисахарид, мономером которого является глюкоза.
Основу бактериальных клеточных стенок составляет вещество муреин (относится к пептидогликанам). У грамположительных бактерий в состав оболочки входят различные кислоты, а сама оболочка плотно прилегает к цитоплазматической мембране. У грамотрицательных бактерий оболочка более тонкая и не прилегает к мембране. Между мембраной и оболочкой образуется периплазматическое пространство. Снаружи клеточная оболочка грамотрицательных прокариот окружена внешней мембраной, составленной из липополисахарида.
У грибов основным веществом клеточных стенок является хитин, а не целлюлоза.
Состав клеточной стенки растений
У растений стенка дочерних клеток образуется уже во время деления родительской. Впоследствии она называется первичной. У многих клеток позже образуется вторичная оболочка.
Первичная клеточная оболочка состоит из микрофибрилл целлюлозы, погруженных в матрикс из других полисахаридов. Отличительной особенностью волокон целлюлозы является их прочность. Молекула целлюлозы представляет собой длинную полисахаридную цепь. Отдельные молекулы соединяются друг с другом водородными связями в пучок, который называется микрофибриллой. Такие фибриллы образуют каркас клеточной стенки.
Матрикс клеточной стенки составляют полисахариды пектины и гемицеллюлозы , а также ряд других веществ (например, белков). Пектиновые вещества представляют собой группу кислых полисахаридов, их молекулы могут быть не только линейными, но и разветвленными. Гемицеллюлозы также смешанная группа полисахаридов. Длина их линейных молекул короче, чем у целлюлозы.
Оболочки соседних клеток растений соединены между собой срединной пластинкой, состоящих из пектатов магния и кальция, для которых характерна клейкость.
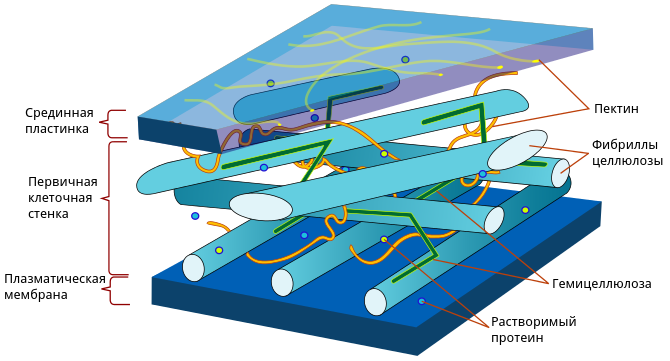
В состав стенок растений входит вода (составляет более половины массы), обуславливая ряд физических и химических свойств полисахаридов.
Жесткий каркас растения во многих местах пронизан каналами (плазмодесмами), по которым цитоплазма одной клетки соединяется с цитоплазмой соседних.
Клетки мезофилла листа (а также некоторые другие) на протяжении всей своей жизни имеют только первичную стенку. У большинства же клеток на первичную оболочку с внутренней стороны отлагается вторичная стенка, составленная из дополнительных слоев целлюлозы. Обычно в это время клетка уже дифференцирована и не растет (исключение составляют, например, клетки колленхимы).
В каждом отдельном слое вторичного утолщения микрофибриллы целлюлозы располагаются под одним углом (параллельно друг другу). Однако разные слои имеют разный угол, что обеспечивает большую прочность.
Часть клеток растений одревесневают (трахеальные элементы ксилемы, склеренхима и др.). В основе этого процесса лежит интенсивная лигнификация стенок (в небольших количествах лигнин есть во всех оболочках). Лигнин не является полисахаридом, а представляет собой сложное полимерное вещество. Отложения лигнина могут иметь различную форму (сплошную, кольцевую, спиральную, сетчатую). Он скрепляет целлюлозу, не дает ей смещаться. Лигнин не только обеспечивает прочность, но и дает дополнительную защиту от неблагоприятных физических и химических факторов.
Функции клеточной стенки
Оболочки разных клеток совместно обеспечивают всему растению и его отдельным частям механическую прочность и опору. Это функция клеточной стенки аналогична одной из функций скелета животных. Однако она не единственная.
Жесткость стенок препятствует растяжению клеток и их разрыву. В результате по физическим законам в клетки может путем осмоса поступать вода. Для травянистых растений тургоцентричность клеток является единственной их опорой.
Микрофибриллы целлюлозы ограничивают рост клеток и определяют их форму. Если микрофибриллы окольцовывают клетку, то она будет расти в длину (поперек направления волокон).
Связанные клеточные стенки образуют апопласт, по которому передвигается вода и минеральные вещества. Плазмодесмы связывают содержимое разных клеток в единую систему — симпласт.
Стенки сосудов ксилемы, трахеид, ситовидных трубок выполняют транспортную функцию.
Наружные клеточные стенки эпидермальных клеток покрыты воском (кутикулой). С одной стороны, он препятствует испарению воды, с другой – проникновению вредных микроорганизмов.
У некоторых растений в определенных клетках оболочки видоизменяются и служат местом запаса питательных веществ.
Структура бактериальной клетки
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядерного аппарата, называемого нуклеоидом. Имеются другие структуры: мезосома, хроматофоры, тилакоиды, вакуоли, включения полисахаридов, жировые капельки, капсула (микрокапсула, слизь), жгутики, пили. Некоторые бактерии способны образовывать споры.
Структуру и морфологию бактерий изучают с помощью различных методов микроскопии: световой, фазово-контрастной, интерференционной, темнопольной, люминесцентной и электронной.
|
Обозначения: 1-гранулы поли-β-оксимасляной кислоты; Клеточная стенкаВ клеточной стенки грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40—90% массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos — стенка). Цитоплазматическая мембранаЦитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм разделены светлым – промежуточным). По структуре она похожа на плазмалемму клеток животных и состоит из двойного слоя фосфолипидов с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты — впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами. ЦитоплазмаЦитоплазма состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул — рибосом, ответственных за синтез (трансляцию) белков. Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Рибосомные РНК (рРНК) – консервативные элементы бактерий («молекулярные часы» эволюции). 16S рРНК входит в состав малой субъединицы рибосом, а 23S рРНК – в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяя оценить степень родства организмов. НуклеоидНуклеоид — эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, замкнутой в кольцо и плотно уложенной наподобие клубка. Ядро бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). Обычно в бактериальной клетке содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК. Капсула, микрокапсула, слизьКапсула – слизистая структура толщиной более 0,2мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка (например, по Бурри-Гинсу), создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например, у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, препятствует фагоцитозу бактерий. Капсула антигенна: антитела против капсулы вызывают ее увеличение (реакция набухания капсулы). ЖгутикиЖгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (1 пара дисков – у грамположительных и 2 пары дисков – у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Жгутики состоят из белка – флагеллина (от flagellum – жгутик); является Н-антигеном. Субъединицы флагеллина закручены в виде спирали. Пили (фимбрии, ворсинки) – нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, то есть за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водносолевой обмен и половые (F-пили), или конъюгационные пили. Пили многочисленны – несколько сотен на клетку. Однако, половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми "мужскими" клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col-плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми "мужскими" сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях. СпорыСпоры – своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий Форма спор может быть овальной, шаровидной; расположение в клетке -терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное – ближе к концу палочки (у возбудителей ботулиэма, газовой гангрены) и центральное (у сибиреязвенной бациллы). Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание. |
