Содержание
• Пили представляют собой внеклеточные белковые структуры, которые осуществляют самые разнообразные функции, включая обмен ДНК адгезию и образование биофильма клетками прокариот
• Многие адгезивные пили собираются с участием системы шаперон-Usher-белок. Сборка происходит на наружной мембране с участием Usher-белка, образующего пору, сквозь которую проходят субъединицы пили, и шаперона периплазмы, способствующего их скручиванию и прохождению через пору
• Жгутики представляют собой внешние структуры клетки, которые служат пропеллерами, обеспечивающими ее движение
• У прокариот жгутики состоят из множественных сегментов, каждый из которых образуется при сборке белковых субъединиц
От поверхности прокариотической клетки отходят два типа придаточных структур, пили и жгутики. Пили представляют собой нитевидные олигомеры белков, присутствующие на клеточной поверхности. Существуют различные типы пилей. Например, F-пили участвуют в клеточной конъюгации и в переносе ДНК. Когда эти придаточные структуры были впервые обнаружены, их назвали «фимбрии» (лат. fimbria — нить, волокно). Их присутствие коррелировало со способностью Е. coli агглютинировать красные кровяные клетки.
Позже для обозначения фибриллярных структур (F-пили), связанных с процессом переноса генетического материала между организмами при конъюгации, был предложен термин пили (или пилюс) (лат. pilus — волос). С тех пор этот термин стал общим для описания всех типов ворсинчатых придаточных структур, и используется наряду с термином фимбрия.
Взаимодействие клеток бактерий с другими прокариотическими и эукариотическими клетками с участием ворсинок часто служит важным этапом заселения эпителия, проникновения микробов в клетки хозяина, обмена ДНК и формирования биопленок. Пили могут служить рецепторами бактериофагов. Основная функция большинства пилей состоит в структурном обеспечении позиционирования специфических молекул, участвующих в клеточной адгезии. Адгезивные субъединицы ворсинок (адгезины) представляют собой минорные компоненты их кончиков, однако основные структурные субъединицы также могут функционировать в качестве адгезинов.
Часто адгезивные пили представляют собой важные факторы заселения микробами организма хозяина. Например, при инфекциях мочевых путей патогенными бактериями Е. coli, клетки прикрепляются к эпителию мочевого пузыря с помощью пилей типа I. Пили этого типа присутствуют у многих грамотрицательных микроорганизмов. Они представляют собой сложные структуры, состоящие из толстого тела, соединенного с тонким фибриллярным концом. На конце расположены молекулы адгезина FimH, которые связываются с остатками маннозы на поверхности клеток хозяина.
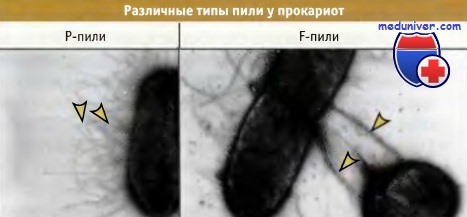 Два типа пили у клеток прокариот.
Два типа пили у клеток прокариот.
Р-пили короче, чем F-пили, и принимают участие в адгезии клеток.
F-пили участвуют в конъюгации и в переносе ДНК между клетками.
Фотографии любезно предоставлены Мэтт Чэпмен (слева) и Роном Скарри (справа), биологический факультет Сиднейский университет.
Сборка пилей представляет собой сложный процесс, в котором участвуют структурные белки, составляющие тело пили, и дополнительные белки, способствующие сборке субъединиц на поверхности клетки. Все структурные компоненты, необходимые для процесса сборки пилей на поверхности грамотрицательных микроорганизмов, должны транслоцироваться через цитоплазматическую мембрану в периплазму и далее, через внешнюю мембрану. В завершении процесса сборки участвуют два специфических белка: шаперон, присутствующий в периплазме, и транспортный белок внешней мембраны, который называется Usher-белок.
Процессы, в которых функционируют эти белки, обеспечивают биогенез более 30 различных типов ворсинчатых структур. Как показано на рисунке ниже, комплексы шаперонов с субъединицами образуются в периплазме и на наружной мембране взаимодействуют с Usher-белком, где высвобождается шаперон. При этом на субъединицах открываются интерактивные поверхности, что обеспечивает их дальнейшую сборку в пили. Исследования пилей типа I и Р показали, что адгезин-шапероновые комплексы (PapDG или FimCH) обладают большим сродством к Usher-белку, и адгезины представляют собой начальные субъединицы, которые собираются в пили.
Включение остальных субъединиц отчасти определяется кинетикой образования на Usher-белке комплекса с шапероном. Наряду с функционированием в качестве сборочной платформы, Usher-белок, вероятно, играет также и другие роли в сборке ворсинок. По данным электронной микросокопии высокого разрешения, PapС Usher имеет вид кольцевых комплексов диаметром 15 нм, которые в середине имеют пору размером 2 нм. После отщепления от шаперона, которое происходит на Usher-белке, субъединицы включаются в растущую структуру пили, которая, как считают, должна выталкиваться через центральную пору комплекса в виде толстой линейной фибриллы, состоящей из одной субъединицы.
Большинство микроорганизмов обладает подвижностью, и часто она обеспечивается длинными структурными придатками, которые называются жгутиками. У грамположительных и грамотрицательных бактерий жгутики собираются на поверхности клеток. Когда на полюсе клетки находится один жгутик, такое расположение называется монотрихиальным (или полярным). Если жгутики расположены вокруг клетки, то такое расположение называется перитрихиальным.
Если на одном полюсе клетки находится группа жгутиков, то говорят об их лофотрихиальном расположении (от латинского «хохолок»). Жгутики бактерий отличаются от этих структур эукариотических клеток, которые состоят из микротрубочек и связанных с ними белков и окружены плазматической мембраной.
Жгутики могут быть различной длины, но их диаметр обычно составляет 20 нм. Они не видны в световом микроскопе, если препараты вначале не обрабатывались реагентами, которые увеличивали диаметр жгутиков. На рисунке ниже видно, что жгутики состоят из трех отдельных доменов: филамента, крючка и базального тела. Филамент жгутика состоит из повторяющихся структур флагеллиновых белков. Флагеллины представляют собой высококонсервативные белки бактерий, что позволяет предполагать, что движение клеток с участием жгутиков характерно для примитивных форм живых организмов. В месте присоединения жгутика к клетке находится базальное тело, представляющее собой сложную структуру, состоящую из множества белков.
Филамент жгутика соединяется с базальным телом посредством крючка. У грамотрицательных бактерий базальное тело проходит через наружную мембрану, протеогликан клеточной стенки и цитоплазматическую мембрану. С наружной мембраной жгутик связан посредством L-кольца. Две пары колец, S-М и Р, способствуют прикреплению жгутика к цитоплазматической мембране и к клеточной стенке соответственно. Каждое кольцо состоит из множества мембранных белков. На цитоплазматической мембране находятся два белка Mot, которые выполняют роль моторов, приводящих жгутики в движение. Еще один набор белков встроен в цитоплазматическую мембрану и выполняет реверсную функцию по отношению к моторам жгутика. Поскольку у грамположительных организмов наружная мембрана отсутствует, у них есть только S-М кольца.
В образовании и сборке филаментов жгутиков участвует несколько десятков различных генов. Их активность строго регулируется в соответствии с порядком процесса сборки. Так, первыми экспрессируются гены, участвующие в сборке базального тела и крючка, а затем наступает очередь генов, ответственных за образование субъединиц жгутика. Экспрессии флагеллиновых субъединиц не происходит до тех пор, пока не завершилась сборка крючка. В этот момент через канал крючка выходит супрессор транскрипции, и, таким образом, снимается подавление экспрессии флагеллина. Субъединицы флагеллина экспортируются через жгутик и добавляются к его растущему концу.
Такой механизм обеспечивает сборку филамента только после образования структуры крючка. Эта структура также имеет отношение к другим секреторным системам белков.
Система хемотаксиса определяет наличие питательных компонентов и затем определяет направление вращения жгутика. В отсутствие питательных компонентов, жгутики вращаются по часовой стрелке, что вызывает поворот клетки. Движение клетки по направлению к молекулам химического соединения или от них называется хемотаксис. В данном разделе мы рассмотрим движение прокариотической клетки в присутствии аттрактанта, являющегося питательным продуктом.
Для того чтобы обеспечить клетке такое движение, жесткий жгутик должен вращаться подобно пропеллеру, за счет энергии, доставляемой протонной движущей силой. Движение клетки состоит из серии прямых пробегов, за которыми следуют ее быстрые беспорядочные повороты. Когда жгутики вращаются против часовой стрелки, клетка перемещается по прямой линии, а при вращении по часовой стрелке клетка совершает повороты. Поскольку в результате поворотов клетка занимает случайные позиции, можно было бы думать, что общий итог движения окажется нулевым. Однако периодичность пробегов регулируется в соответствии с доступностью питательного компонента: более длинные пробеги характерны для движения клетки по направлению к источнику питания, и количество поворотов возрастает, когда клетка направляется от него.
Хотя направление отдельных пробегов все еще случайно, общий результат проявляется в движении клетки в сторону аттрактанта.
Пути передачи сигнала хемотаксиса у прокариот характеризуются чрезвычайно консервативной природой. Единственным из известных организмов, в геноме которого отсутствуют гены хемотаксиса, является Mycoplasma. Практически у всех прокариот обнаружены следующие консервативные белки хемотаксиса: CheR, CheA, CheY, CheW, и CheB. При протекании сложного каскада событий, включающих фосфорилирование и метилирование, эти белки обеспечивают сложный, скоординированный и высокогибкий ответ клетки на присутствие аттрактантов и репеллентов в окружающей среде. Мы опишем, как происходят эти события в клетках Е. coli.
Присутствующие в окружающей среде аттрактанты или репелленты связываются с рецепторами, расположенными на цитоплазматической мембране. С этими рецепторами взаимодействует киназа CheA, также расположенная в цитоплазматической мембране. Эта киназа фосфорилирует CheY, который затем связывается с мотором жгутика, что приводит к переключению направления его вращения и к повороту клетки. Под действием фосфатазы CheZ из CheY удаляется фосфатная группа. При низкой концентрации аттрактанта происходит аутофосфорилирование CheA, фосфатная группа переносится на CheY, и последний мигрирует к мотору жгутика, изменяя характер движения клетки на поворот.
Система хемотаксиса характеризуется еще одним уровнем сложности, который позволяет клетке постоянно адаптироваться к условиям, существующим в окружающей среде. По мере своего продвижения по градиенту концентрации химических соединений, клетка может реагировать на возникающие небольшие флук-туации. Такая кратковременная память обеспечивается за счет метилирования мембранных рецепторов. CheR метилирует мембранные рецепторы, a CheB удаляет метальные группы.
Метилирование рецепторов увеличивает активность киназы CheA, что приводит к десенсибилизации системы. В свою очередь, CheB также фосфорилируется CheA; это вызывает увеличение метилэстеразной активности CheB и замыкает цикл обратной связи для сигнального каскада.
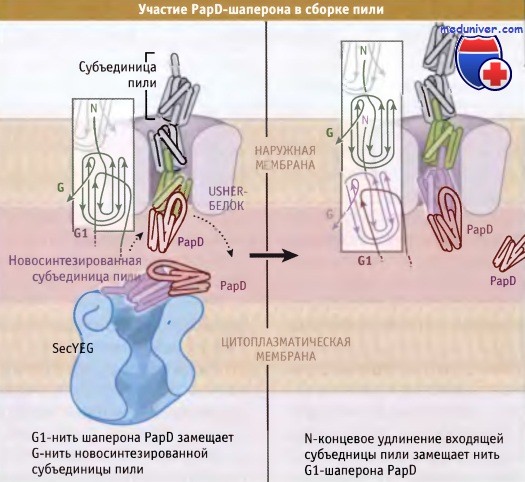 Для сборки пили необходим шаперон PapD и мембранный Usher-белок.
Для сборки пили необходим шаперон PapD и мембранный Usher-белок.
Слева: нить G1 шаперона PapD завершает формирование нативной структуры новосинтезированной субъединицы пили,
располагаясь между двумя нитями этой субъединицы.
Справа: в процессе сборки пили N-концевая нить новосинтезированной субъединицы замещает G1-цепь шаперона PapD,
связанную с предыдущей субъединицей пили. 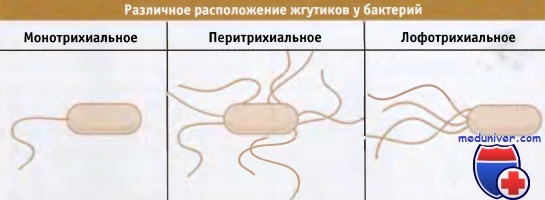 У различных типов бактерий жгутики расположены по-разному.
У различных типов бактерий жгутики расположены по-разному. 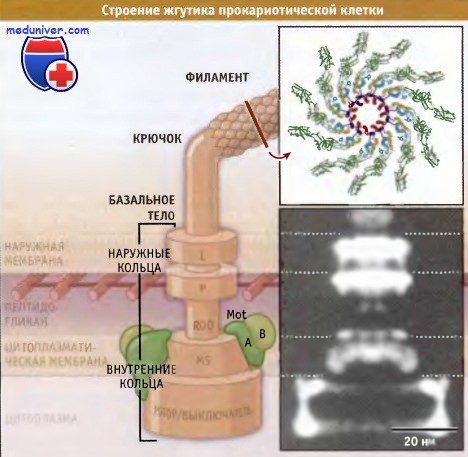 Жгутики прокариот состоят из трех сегментов, каждый из которых содержит многочисленные белки.
Жгутики прокариот состоят из трех сегментов, каждый из которых содержит многочисленные белки.
На врезке вверху представлено изображение структуры жгутика, полученное с помощью электронного микроскопа,
а внизу — электронная микрофотография области базального тела и крючка.
Филамент жгутика состоит из флагеллиновых субъединиц, которые собраны в спиральную структуру, содержащую 11 субъединиц на виток. 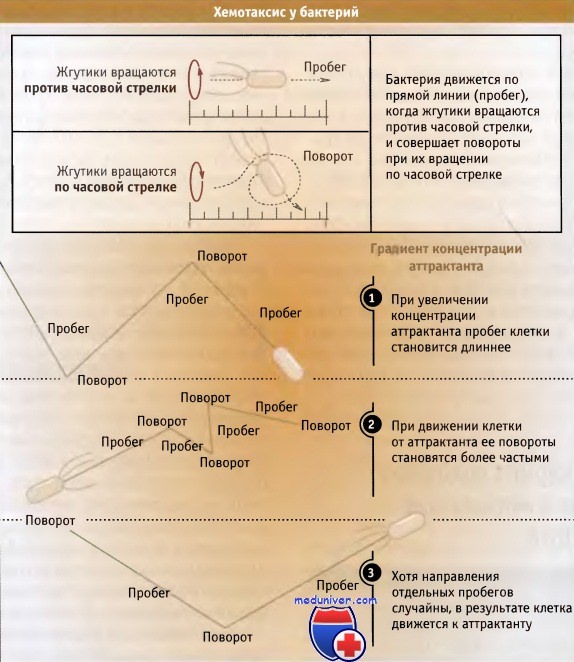 Движение бактерии по направлению к молекулам питательных веществ
Движение бактерии по направлению к молекулам питательных веществ
и от них можно представить в виде длительных прямолинейных треков и, соответственно, большим числом поворотов.

Пили́, или фи́мбрии, или ворси́нки [1] — нитевидные белковые структуры, расположенные на поверхности клеток многих бактерий. Размер пилей варьирует от долей мкм до более чем 20 мкм в длину и 2—11 нм в диаметре. Пили участвуют в передаче генетического материала между бактериальными клетками (конъюгация), прикреплении бактерий к субстрату и другим клеткам, отвечают за адаптацию организмов, служат местами прикрепления многих бактериофагов.
Содержание
Структура [ править | править код ]
Пили состоят из одного или нескольких типов спирально уложенных белковых молекул, которые называют пилинами [en] или фимбринами. Часто на концах пилей присутствуют особые прикрепительные субъединицы (адгезины), но иногда весь пиль обладает адгезивными свойствами. Структурно пили могут быть от тонких нитевидных образований до толстых палочкообразных структур с осевыми отверстиями. Очень тонкие пили (диаметром менее 2 нм) называют «кудряшками», они сливаются в пушистую липкую массу на поверхности клеток, которая обеспечивает их агрегацию. Пили-кудряшки являются амилоидными [en] фибриллами, родственными белкам, формирующим амилоидные бляшки в мозге пациентов с болезнью Альцгеймера [2] . Часто пили разбросаны по всей поверхности клетки, но иногда локализуются только на одном её конце [3] .
Функции [ править | править код ]
Основная функция большинства пилей заключается в обеспечении правильного позиционирования особых молекул, обеспечивающих адгезию клеток. Взаимодействие бактерий с другими про- и эукариотическими клетками, обеспечиваемое пилями, является важным этапом заселения эпителия и проникновения патогенных бактерий в клетки организма хозяина, образования биоплёнок и переноса генетического материала при конъюгации. Некоторые пили задействованы в движении бактериальных клеток. Пили иногда служат рецепторами бактериофагов. Например, при инфекции мочевыводящих путей [en] , вызванной патогенными штаммами Escherichia coli, клетки прикрепляются к эпителию мочевого пузыря с помощью пилей, на конце которых находятся молекулы белка-адгезина FimH, который взаимодействует с остатками маннозы на поверхности эпителиальных клеток. Благодаря прикреплению к эпителию клетки бактерии не выводятся из организма с мочой. Пили являются важными факторами вирулентности [en] многих кишечных бактерий, таких как Salmonella enterica, энтеропатогенные штаммы E. coli [en] , Vibrio cholerae. Прикрепление пилей к эукариотическим клеткам может запускать в них определённые сигнальные каскады. Например, прикрепление пилей Neisseria к эпителиальным клеткам вызывает в последних выход ионов кальция в цитоплазму, при этом кальций является важным вторичным посредником в передаче сигнала в эукариотических клетках [4] [5] .
Образование [ править | править код ]
В образовании пилей, помимо самих белков-пилинов, участвуют дополнительные белки, способствующие правильной сборке. У грамотрицательных бактерий они должны пройти через клеточную мембрану, периплазматическое пространство и наружную мембрану. Ключевую роль в сборке пилей играет периплазматический шаперон PapD и транспортный белок внешней мембраны, известный как Usher-белок. PapD связывается с пилинами, вышедшими в периплазму, и доставляет их к Usher-белку, после чего высвобождается назад в периплазму. Доставленные к Usher-белку пилины приобретают активированную конформацию и далее собираются в пили [6] .
Пили-кудряшки [en] образуются по специфическому, тщательно регулируемому механизму. У E. coli в их сборке задействованы белковые продукты оперонов csgBA и csgDEFG. Фибрилла кудряшки образована белком CsgA, который, наряду с минорным белком CsgB, участвует в фазе нуклеации фибриллы. Когда нуклеация пройдена и начинается рост ворсинки, CsgB включается в неё. Фибриллы CsgA очень устойчивы и разрушаются лишь под действием 75 %-ной муравьиной кислоты. Оперон csgDEFG кодирует транскрипционный фактор CsgD и три предполагаемых фактора сборки пилей [2] .
Классификация [ править | править код ]
Традиционно пили подразделяют на 4 типа.
- Пили I типа связываются с остатками маннозы на других клетках;
- Пили II типа независимы от маннозы и не вызывают гемагглютинацию;
- Пили III типа вызывают агглюцинациюэритроцитовчеловека, обработанных танниновой кислотой;
- Пили IV типа также известны как половые пили, они участвуют в процессе конъюгации [7] .
Некоторые учёные подразделяют пили на пили главного типа и альтернативного типа. Пили главного типа включают пили типов I—III по традиционной классификации. Они располагаются по всей поверхности клетки и прикрепляются непосредственно к её наружной поверхности. Пили главного типа выполняют только адгезионные функции. Пили альтернативного типа (тип IV по традиционной классификации) образуются в количестве 1—2 на клетку и располагаются на одном или двух её полюсах. Они, подобно жгутикам, имеют базальное тело и заякорены в клеточной мембране, а потому насквозь проходят периплазму и наружную мембрану. Пили альтернативного типа участвуют не только в адгезии. Они могут быстро разбираться от основания, при этом дистальные их концы остаются прикреплёнными к субстрату или другой клетке. За счёт этого одна клетка может подтягиваться ближе к другой, что особенно важно при конъюгации [8] .
Микробиология
Предметом изучения микробиологии

Жгутики и пили бактерий. Электронная микроскопия. (Атлас по медицинской микробиологии, вирусологии и иммунологии / Под ред. А.А. Воробьева, А.С. Быкова – М.: Медицинское информационное агентство, 2003.-236 с.).
Escherichia coli. Электронная микроскопия. Адсорбция фага ms2 на f-пили. х100000.«Авакян А.А., Кац Л.Н., Павлова И.Б. Атлас анатомии бактерий, патогенных для человека и животных. М «Медицина».-1972.-183 с.»
“>
, – тонкие полые нити белковой природы, более тонкие и короткие (3-10 нм х 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина. Они обладают антигенной активностью. По своему функциональному назначению пили подразделяются на несколько типов.</p>
<p>Пили 1-го типа, или общего типа – common pili – пили, ответственные за адгезию, т.е. за прикрепление бактерий к поражаемой клетке. Они начинаются на ЦПМ и пронизывают клеточную стенку. Их количество велико – от нескольких сотен до нескольких тысяч на одну бактериальную клетку. Бактериальные и эукариотические клетки заряжены отрицательно, но поверхностные микроворсинки снижают заряд бактерии и уменьшают электростатические силы отталкивания. Кроме того, увеличение площади поверхности бактериальной клетки дает ей дополнительные преимущества в утилизации питательных веществ окружающей среды.</p>
<p>Пили 2 типа (половые, F-пили, конъюгативные – sex pili) участвуют в конъюгации бактерий, обеспечивающей перенос части генетического материала от донорской клетке к реципиентной. Они имеются только у бактерий-доноров в ограниченном количестве (1-4 на клетку), более длинные (0,5-10 мкм). Отличительной особенностью половых пилей является взаимодействие с особыми «мужскими» сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.</p>
<p style=)
